
| THE INTEGRITY PAPERS | Falk - Genre Group | ceptualinstitute.com |
Background: some basics of biology
To understand why I am so enthusiastic about Dean Falk's research dealing with humanity's deep ancestral relatives, it is necessary for me to take you on a survey journey of understanding - about how biological systems are generally organized, and how they function among a variety of different but subtly interconnected levels.
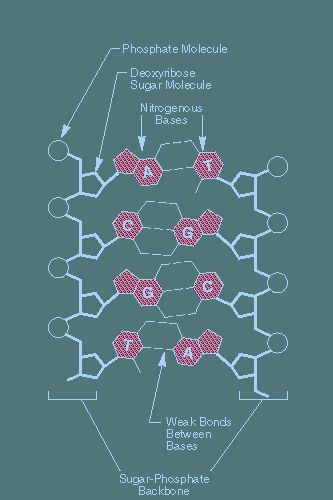
The letters "DNA" have been heard of - even if not perfectly understood - by most people of tolerable education and news exposure. DNA and RNA are long chains of organic molecules formed from a family of sugars and organic acid molecules (4 main ones) which are very sturdy and robust. Not extraordinary in and of themselves, these 'amino acids' easily link with another molecule in their own group, in very specific pairs meeting at a certain end location of their atoms and trailing off in opposite directions - like two bar magnets that comfortably join end to end. One double-set uses two bonding links and the other, three links. They are very pair specific this way.
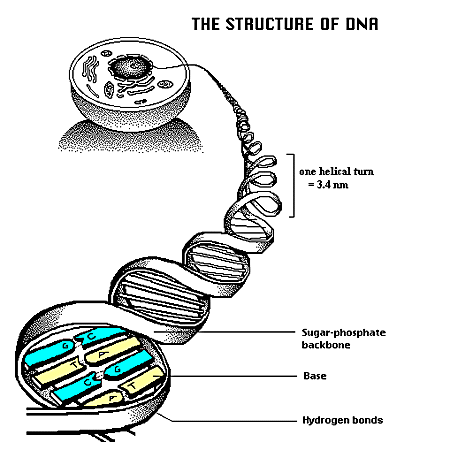
The next arrangement that occurs is a stacking of these fairly flat molecule pairs into layers. This is done by Phosphate bonds between the outer edge sugars, which form the scaffolding ... two outer edge chains that backbone the inner amino acid 'information' molecules, lined up together in long sequences. In order to stack - with the least strain on this 'layering' of their atoms - the molecule pairs join with a staggered offset, pivoting around those central 2 and 3 point bonds. The result is a spiraling of these layers into what we now know of as the "double helix" formation of the so-called "genetic material" of living biological organisms. Residing deep inside the protected nucleolus (nucleus of the nucleus) of every living cell, these long chains of molecules act like a primary instruction manual which can chemically interact with other biological chemical pieces in their soupy fluid cellular environment, and so, direct activities out in that environment.
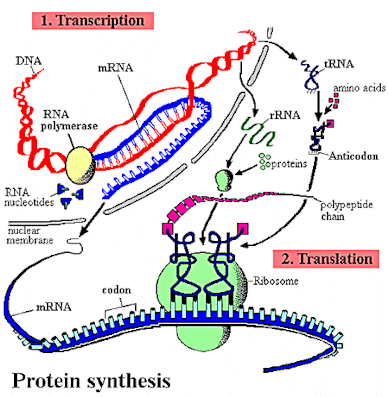
Those inner double and triple bonds are relatively weak and can un-bind much like a zipper. Sometimes when this happens, other loose amino acids rush in and fill in with the zippered-open strands. This is the essential act of cell division/duplication/multiplication. Most other times, lengths of open strands of slightly different 'RNA' (DNA, but with added oxygen atoms) find their way near and outside the nucleolus and are involved with this binding/unbinding process. When this happens, sequences of RNA go out into the mix of other chemicals and molecules, both as mirror copies of original DNA strand lengths, and as double-mirrored copies (which then duplicate the original sequences exactly).
There, new types of molecules in the cytoplasm (soup of cell fluids, chemicals) bind to the RNA. These connect to the RNA and themselves also and form yet other long strands of important stable working molecules. These become what we call proteins, enzymes, and other metabolically important molecules, which then go off to interact among themselves elsewhere. By how their binding atoms link up with yet other molecules, or, by how they accept and then hand off flows of electrons of different energies, they directly build the physical parts of what make a cell ... mitochondria, walls, etc.... or participate in the processes that keep a cell energized and alive, taking in outside environment molecules to be used as food and passing off unusable or energy drained molecules.
With slight variations allowed, this process is generally known as the 'central dogma' of protein synthesis.

To re-cap this scenario: Atoms form molecules. Molecules of certain special forms (DNA/RNA) then do things that single atoms couldn't. A menagerie of other simple molecules then become organized by the patterns of DNA/RNA and go on to do other things that the DNA/RNA couldn't do themselves. They interact, form, and generally "behave" in new ways and with new skills. The seemingly noisy randomness of electrons jumping in tiny energy amounts inside single atoms and among a few atoms that connect together, first become one pattern of coordinated organized energy structures and "flows", and then help cause the formation and activity of yet others. Different, new, competent. A brick by itself is interesting. A group of them can form a pile. On top of that can go a cover. But a pattern of bricks can form a wall. And several walls can stand near each other. So when that cover is placed, it now becomes a roof, and the random bricks are now a "house". A brick is not a house, yet without bricks, this kind of house cannot be. Coordinated connections produce new qualities. And all sorts of wonderful things, abilities and behaviors emerge and blossom from .... "communication" ... the ways that things and energy connect with each other.
Assemblies of assemblies (Hebb, 1949)
After this, in their turn, cells come into being with many different functions, abilities and forms. Some are suited for message transmitting, some for shrinking and extending in unison like trillions of coordinated in-line pistons, some do other things. The first group becomes the basics of neuron cells - brains, eyes, inner ears, etc., capable of sensitivity to different kinds of electromagnetic and light and sound and taste and feel energy. The second group becomes the many kind of muscles -- large ones like those connected to bones and helping the whole body of an organism move with conscious intention, and continuing down to very minute ones that wrap into tubules - arteries, veins and capillaries that move blood with all the aeration and nutrition which that fluid can carry. And, of course, there are other cells types which function and form in myriad alternative ways.
As far as where all-important 'energy' comes from, there is only one source - which energizes these phenomena via two pathways. This is a very important idea to grasp...the double pathway ( 1972, 1999). The bulk of the energy which powers living systems originates in its usable form inside the electron clouds and layers of each individual atom. It is here where the fundamental relationships of 'universe' produce the wellspring of energy-in-motion. When one of the primal units of universe - light - comes into being it travels with one speed: "fast". As atoms organize from smaller components, there are several 'fundamental forces' which coordinate with each other and with this "speed of light" factor, to produce 'atoms' which have inherent energy - contained in the way electrons can and may move in kinds of 'orbits' around an atom's central mass of other atomic particles.
The first path of the energy of life comes from these 'spontaneous' motions and energies of the electron clouds, of all the different possible atoms. There are rules to how they all work, but that you can learn about elsewhere. The main thing to recognize is that the outer edge electrons of atoms are allowed to exist in different energy levels, which depend on the light energies they contain. Electrons tend to move in paths comfortable with the amount of energy they carry. When atoms meet, these "always energized" electrons may encounter pathways that allow them to move around several atoms very comfortably. Again, the specific processes are available in any good physics book. Just remember, compatible electron pathways and energy levels which can adjust and coordinate together, make up the essential first path of what enables atoms to form molecules.
The second path of the energy of life happens when so many atoms congregate that the force of gravity overwhelms the electromagnetic interactions described above. The result of this second path is: stars.
Huge accumulations of atoms force electron energies to come together in highly stressed ways that squeeze into small spaces and into each other. As these energies mix and bind and break in every higher amounts violent energy releases occur. It is this form of released light and electrons - from our star, the sun - that bathes over and through the slower and 'cooler' atoms and molecules we spoke of earlier. This flood of energy helps move the interactions of molecules along, encouraging the total processes of life.
One source, two paths. One source, two paths.
Sunlight provides the extra energy and nutrition that comes into these many-formed systems. It prompts all the bits and pieces and processes that connect and interconnect, unconnect and reconnect, build, breakdown and re-build again. And all this 'internal' communication activity is mirrored on the even larger scales by all sorts of external interaction communications. Involvements with companion other life forms. Involvements with companion same life forms. This is nature, society and the relations of life. As we know from experience, existence doesn't include just one form of living organism. It includes so many that it can be mind boggling to consciously think of them all - but not impossible(!). And the awe inspiring enormity of it all can't stop us from recognizing the uniformity and shared processes.
Thinking of our original descriptions of DNA plus, what does this all mean? It means that four simple molecules, depending on which ones are next to others - in long strings of possible arrangements - have ended up producing all manner of different next-step molecules that interact together in, again, different ways to produce the terrifically large possibility of tissue and body shapes and forms and working-together connections of all the wild and infinite diversity of what life can look like, can be and can behave as.
Imagine then, what this deeply means if we think in terms of the next-step next-composition steps! In all of life there are physiologies - healthy working interaffective metabolisms of all manner of chemicals and such - all doing pretty much the same general processes (like "breathing", processing "food") yet doing them with slightly different molecules and in different combinations, and yet, all working well together in their own ways and together. They produce organisms that do different things, react and act in alternative ways, because the inner instructions are different, producing a different chemical soup of molecules, that combine in alternative ways and produce the special physical bodies which are capable of special actions, and thus, special ranges of behaviors.
Animals with gills can process aeration gasses (like oxygen) saturated in fluids. Animals with lungs can do it totally gaseous environments. Animals with appendages called wings can fly by themselves (well, most can), but animals without wings cannot.
From these observations, it's an easy connection for us to think about strings of DNA that produce proteins and so on, that organize into hollow bones and to robust muscles and to structures called feathers that, all together, enable an animal called "bird" to perform the act of flying. Like an architect's blueprint, we can envision the inner genetic code calling for the material that can form just so - in size, dimensions, twists and turns of form and structure - and so produce some absolute body form that can function in some absolute way, and show some characteristic, like wings, like eyes colored blue.
But what if the process is not limited to direct genetic coding for form per se? Might there be something more subtle at work? Something so intricate and ingenious that we've missed appreciating the extent of what genetic material really dynamically accomplishes and the full scope of how it works?
I strongly think so. And it all has to do with how we comprehend the extent and quality of 'complexity' in the world. Where I'm going here is not into the mechanics of metabolism but into the impacts that small and subtle changes can have, and so go on to produce large or unusual ability-effects someplace else. Effects that we indeed tend to place a lot of special value on, as if they were the important tangible qualities of life, yet having no identifiable origin within the genetic code.
My message to you is that the importance of genetic material is not just its ability to harbor consistent information that produces viable instructions for making other viable assemblages of behaving systems, but in its capacity to create and transfer information can cascade and blossom when it reaches those other levels, producing special and extraordinary events, all from the merest of minimal "inconsequential" alterations.
What I glean from Dean Falk's work is the possibility of a clear example where this can be pinpointed as actually having happened. It is not proven fact, it is my hypothesis. But it is a scenario of true complexity dynamics - one based on solid established principles and researched understandings - that conjectures a plausible way to integrate the diversity of genetic and physical events, and see them as a real thriving complex whole. In the most accurate and extended sense, not a local or limited mathematical one.
The focal question is whether - in some instances - the genetic code is an "absolute" blueprint or a "relational" one. Here, whether the code specifies the shape and mapping of a blood circulatory system directly - "grow a certain length, bend 80 degrees, grow some more, bend back 100 degrees, continue to grow and branch in a set predetermined pattern", or, indirectly - yet producing the same end result systemic pattern. And if "indirect", how subtle then could such an initial genetic event be? And if "indirect", what could be the result? How un-subtle? How truly significant? What could it mean for human existence, and indeed, for how we are in the world we perceive and persist in?
******
From "Braindance", Dean Falk. 1992. Henry Holt and Co., New York.
The Evidence from Cranial Blood Flow
p.132 .... "Then in 1982, quite by accident, I found myself immersed ...."
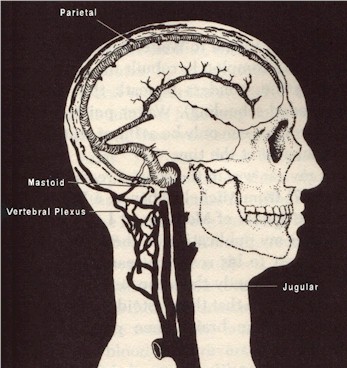
"Cranial remains of australopithecines and early Homo were my subject matter and, once again, I was spending my summer doing research in African museums. For the first time, I also saw numerous casts of the cranial remains from the Hadar early hominids. Although convolutions and sulci were my primary interest, that summer I became fascinated with a particular feature that showed up on some of the fossils. It had to do with the cranial blood flow."
"The feature that caught my attention was a dramatic groove located inside the back of the braincase. This large groove is called the occipital/marginal (or O/M ) sinus, and it is rarely present in a form large enough to leave a groove in apes or living people. In life, the enlarged O/M sinus of early hominids provided a major pathway for draining blood from the skull. But only for some early hominids. What intrigued me was that all of the scorable robust australopithecines (there are now seven) and all of the scorable Hadar early hominids (five specimens) had this feature, while only one of the (now) six gracile australopithecines showed it. 10 (That specimen was Taung, which may have been in correctly placed with gracile instead of robust australopithecines because of its young age, as the reader will recall.) Contrary to Johanson and White's taxonomic scheme in which robust australo- pithecines and the Hadar early hominids were completely separated, presence of the unusual O/M sinus in all scorable members of both of these two groups suggested to me that they might, in fact, be directly related".
"But why did some early hominds have an O/M sinus? What was its function? For that, I turned to my colleague and fellow paleontologist, Glenn Conroy, now in the Department of Anatomy and Neurobiology at Washington University School of Medicine. Glenn agreed that it seemed to be too much of a coincidence to find the highly unusual O/M sinus in all members of two different groups unrelated early hominids. We decided to put our heads together about heads."
"Glenn and I knew that Lucy {CI memo: "Lucy" is the nickname given the earliest recognized likely evolutionary ancestor of humans} and company were among the earliest known bipeds. That led us to wonder if the O/M sinus had something to do with the origin of bipedalism. As a starting point, we decided to find out whether or not there was a general relationship between posture and cranial blood flow in living people. Within weeks, we located and studied a number of relevant clinical and physiological reports that made it clear that cranial blood flow depends very much on posture. And not just for humans."
"Monkeys, snakes, people, and giraffes are all subject to the constraints of gravity. In all of these animals, blood flow changes with a shift in body posture from a horizontal (lying down) to a vertical (standing up) position. (For snakes, cranial blood flow differs between horizontal positions on the ground and vertical hanging posture in trees.) A major reason why blood flow changes with altered postures is because gravity places different stresses (called hydrostatic pressures) on horizontal and vertical columns of fluid (such as blood in veins)."
"To understand this, imagine a long tube half filled with water. If the tube is vertical, all of the water is in its bottom half. When the tube is completely tilted over on its side, however, the water is redistributed along its entire length. Now imagine that the tube contains little holes that are sparsely but evenly distributed over its entire surface. When the tube is vertical, the water drains out of the holes in its bottom. When it is horizontal, on the other hand, the water exits through holes all along the tube's side. To wit: Different postures, different exits for fluid. And all because of gravity."
"Conroy and I learned that the same thing happens with blood that is draining from peoples' skulls. If an individual is lying down, cranial blood flows out through the internal jugular veins. When the person stands up, however, most of the exiting cranial blood shifts away from the jugulars and into a network of veins that surrounds the spinalcord. That network of veins is called the vertebral plexus. Our bipedal existence depends on it! We were on to something."
"Our next task was to determine if there was a relationship between and enlarged O/M sinus and the vertebral plexus of veins. Again, we studied the literature and , again, the answer was affirmative. Most humans have small O/M sinuses (not big enough to leave grooves in skulls) and these usually have numerous connections with the vertebral plexus of veins. More to the point, in a study of one hundred human cadavers, nine had relatively large O/M sinuses.11 Seven of these nine O/M sinuses were connected to the vertebral plexus of veins. Clearly, it was a good bet that the even larger O/M sinuses of early hominids were also connected to the vertebral plexus of veins. After all, if cranial blood flows to the vertebral plexus of veins when a person stands up, it makes sense that a route leading to the vertebral plexus would be enlarged in some of the earliest bipeds."(p.135) . . .
. . .(p.152) "The solution that Conroy and I proposed to explain the function of O/M sinuses in certain groups of early hominids raised another question. If O/M sinuses were the product of selection for bipedalism in early hominids, as we claimed, why don't bipeds like ourselves have them in higher frequencies?" . . . "..when humans stand up, the blood leaving their skulls shifts away from the jugular veins into the vertebral plexus of veins that surrounds their spinal cords. ... The question now became: How do living people deliver blood to their vertebral plexus of veins when they are upright, since their O/M sinuses are usually so small?" ..
. . . "In other words robust australopithecines and the Hadar early hominids had one enlarged route that could shift blood to the vertebral plexus when they stood up, but modern people have a more diverse, complicated network ... to carry out this function." . . "(Apparently) different systems for cranial blood flow had evolved in different groups."
"...could this hypothesis be tested? .... in the fossil record, it could. ... A few of the many little veins that contribute to cranial blood flow in living people go through holes (foramina) in the skull. The particular veins are called emissary veins and they have names like mastoid (near the back of the ear) and parietal (at the top rear of the skull) emissary veins. Emissary veins don't exist for most cranial blood veins, but the few that do exist provide a glimpse into the entire system. ... the foramina through which emissary veins travel are often preserved in fossilized skulls."
"I decided to spend the summer of 1984 counting these little holes in skulls in museums in Africa and Europe. .. I did. Lots of holes in lots of specimens. Whole skulls were examined from 50 chimpanzees, 50 gorillas, and 50 modern humans. In addition, I scored every fossil cranial fragment I could get my hands on for gracile australopithecines, robust australopithecines, Homo habilis, Homo erectus, archaic Homo sapiens, and Neandertals. ... relevant data that could be found in the literature were added ..and included in my analysis."
..."Interpreting these data .. it appeared that increased frequencies of emissary veins were ... accompanied by increased frequencies of other non-traceable blood vessels. ... the whole system of cranial blood vessels had undergone a dramatic evolution, becoming more complex in hominids leading to Homo sapiens ..."
... "Of Car Radiators and Letters from France"
"... I received a letter (dated January 13, 1987) from Michel Cabanac, a French physiologist who is now at the School of Medicine at Laval University in Quebec." {Ceptual Institute 1998 references: Cabanac '91, '92, '97} " ...my publication on the evolution of emissary veins was of interest .. (since) Cabanac's group had shown that the mastoid and parietal emissary veins help to cool the brain when people are overheating."
"The brain is an exquisitely heat-sensitive organ. ..."
"Whereas I had .. speculated that the mastoid and parietal emissary veins provided windows into a wider, more complicated network of cranial veins, Cabanac and Brinnel had actually demonstrated this to be the case." ..
"Cabanac and his colleague Heiner Brinnel .. (demonstrated) ..that the mastoid and parietal emissary veins provide (proof of) more complicated network of cranial veins .. Cabanac and Brinnel .. concluded that when blood flows into the braincase in the ophthalmic, mastoid and parietal veins, it also flows inward in the innumerable other veins that comprise the cranial network." ...
"Cabanac's letter had a profound influence on me. In a key portion, he wrote:
'It is possible that emissary veins were developed for the defense of brain temperature. Brain size has increased with bipedalism. The brain's thermolytic (cooling) needs have increased with its increasing size. ... This hypothesis does not contradict yours but might complement it. I would be interested in hearing from you.' ...
The impact of Cabanac's research hit me in the middle of the night. ... I got up ... and went to the lab to plot hominid cranial capacity against the data for emissary veins. ...
It was beautiful. For the past two million years, the increase in frequencies of emissary foramina kept exact pace with the sharp increase in brain size in Homo. Clearly, the brains and veins had evolved rapidly and together.
To capture the importance of what you've just read, consider this: A shift in how blood drains out of the brain and skull case impacts the body's ability to better or worse control its temperature. A shift in how blood drains out of the brain and skull case impacts the maintenance of blood pressure to the most vital organ, making it easy or difficult to continue vision, speed of response to critical events, sense of balance and coordination, and thousand of other subtle abilities, which then filter into how an animal can act and behave in its environment and with its companions. A re-routing of blood flow path isn't just a matter of alternatives - convenience in getting from 'here' to 'there' - it impacts very intimately what an animal can or cannot do ... the intangible behaviors that are more of our hallmark of what a living thing 'is', than its physical form per se. A shift in how blood drains out of the brain and skull case impacts the stamina and long-term performance abilities of a creature and can have a profound effect in what behaviors it and its companions are capable of. It can open vistas of behavioral oppor- tunities that can distinguish them from their predecessors. A simple shift in the path of how blood drains out of the brain and skull case can do this. ***** My exposure to Dean Falk's work came in 1996, when I caught a brief glimpse of her interview on the Discovery Channel. The profound importance of her discovery resonated with me immediately. The science of Complexity's view had been heading in the direction that mathematical relationships underscore all sudden arisings of 'new phenomena". The core foundation of 'traditional' Complexity is fractal mathematics. Their premise is that previously unobvious patternings and dynamics wondrously "emerge" - totally unpredicted - from special relational equations. These mathematically novel functions - found to have several models in real physical systems (like certain heart-beat rhythm patterns) - were then generalized. It was assumed that all novel-complexities must be the result of fractal and related functions.With the Integrity Paradigm, I was going in an alternative and more general direction that that. And Falk's discovery provides an exemplary empirical - real physical - example, that I can point to in support.
One essence of Integrity dynamics says:
Behaviors at any one level of organization in a system correspond with
other important behaviors elsewhere in the next larger and next lower
levels of assemblies; in local and non-local ways.
The important dynamic is:
Open communication both within-levels and among nested-levels
of architecture and performance.Now, something very special happened those many millions of years ago. Something which altered the physical architecture of blood circulation paths in an early hominid line. It happened in conjunction with growing a bigger brain and with other physical changes that were allowing the creatures to walk more upright as a matter of daily routine. Slightly improved visual vantage points. Hands freed from the task of locomotion. Improved ability to endure warmer climates or terrains. Physiologically improved control of blood pressure. Longer waking hours possible. These and many more subtle changes which we can surmise, encouraged profound changes in behaviors - both for individuals and for their collective social groups.
How did this all come about? What orchestration of serendipity brought the necessary aspects together? Could the standard model of DNA - the repository of coded information which inform cells what they are and so what functions they are able to perform - accomplish this orchestration?
The current "mapping model" of DNA genetics holds forth that certain strings of amino acids - called "genomes" - code very specifically for physical structures, and therefore, for functions. There are codes for arm bones, for eye color, for heart, for neurons, for ... the blood circulatory system. Code here, structure there. As 'proof' is the simple observation that we can publish a book on human anatomy. What we identify as present in one body, will be present (with minor variation) in any other. We rely on this reality of fact and relation. We assume that DNA codes specifically for 'behaviors' as well as physical structures.
But, carried to its extreme, could it be that so many millions of years ago, a gene code suddenly went from "vein, grow relatively straight downward" to "vein, make a sharp right angle turn, grow, turn the opposite way, keep growing"? I mean, if genomes code only for specific structure, then if we observe a different structure, there must be a genome that specifies that structure. As an aside, Fractal Complexity theorists might say, "yes". Their version would be: well, all fractal functions have two or more 'attractor basins' ... that is, there is a tendency to choose one of several potential ways of settling-in, so to speak. The growth patterns probably represent two likely 'basins' (formation possibilities), and something happened in the genetics which 'flipped-the-switch' and allowed the same basic genetics to produce its 'alternative' state ... whether the alternative was originally apparent or not.
But if that were the case, regressive - devolving - species should continually appear along side evolving ones. An 'attractor basin' approach to species development would require that any new species could just as easily 'evolve' into its predecessor species. We would see continual examples of flip-flopping speciation in every new generation of animals. Fortunately, we do not see this process. (Now, there are environmentally pressured 'regressions' of species that occur over long time spans, such as once landed-based mammals which have returned to marine environments, like whales and dolphins, but that is another process.)
Consider now that we have to account for this shift in vein paths, but without having to have every centimeter twist turn and change coded for in the DNA. Is it possible? Yes.
Keep in mind a few things. In a developing fetal body all the tissue is soft and flexible, even bones. We have a situation which I characterize as "vascularization before calcification". Simply: soft tissue development precedes rigid tissue hardening. This means, for example, that both blood vessels and skull bones are forming at the same time. Skull bones will harden around blood vessels which happen to be in an area, rather than vessels forcing their way through hardened bone. The foramina which Falk and Cabanac/Brinnel were concerned with must have formed after the veins were already present.
The issue now becomes: What caused the relocation of vein paths? Do we have to rely on a DNA code that specified the exact turns and alterations (with emphasis on 'exact')? No.
Rather, the scenario goes something like this. Inside the developing hominid brain/skull other tissues were growing and competing for space. Brain matter itself, as well as the blood system to nurture it. Now, the way DNA works is that there are special short sequences which act as stop-flags. What ever process is going on and information sequences being copied for enactment, at some point the stop-code is reached and other activity is started.
Now suppose that the sequence which encouraged continued brain tissue growth was started just a little earlier than the sequence for circulatory tissue. Or, was allowed to continue longer because the stop-code was shifted to further along the sequence. Or more growth protein was infused into that area of brain growth. (There are several alternative, but equally plausible explanations, none which can be confirmed with absolute accuracy, but which can be conjecture based on known laboratory observations.) What would be the result?
In the first place, added brain tissue growing in a relatively confined region would compete with other tissue just for physical placement. In this scenario, the additional brain matter pressing into an area would displace the tissue around it. In this case, veins of the circulatory system. So what do we have as a result? We don't have genes that necessarily code for veins to turn here, grow there. What we do have is a slight change in the gene sequence that codes for how long certain proteins are produced - in ways different than done in past members - and that change of production-time causing physical changes in tissues that it doesn't directly control. (!)
What we have is a subtle change in the metabolic chemistry that produces a visually distinguishable change in the physical form of adjacent companion tissue - not its own tissue directly - and that adjacent change (vein path) coordinating with the original one (more brain mass) to produce profound profound changes in the body structure, the body functions, the body abilities, the body behaviors, and the creation of new lineages of life forms.
This is the Integrity view of full and extraordinary complexity in our universe. We are a concordance of behavior spaces communicating with other behavior spaces in such exquisite ways that the grandeur of existence is at once powerful and delicate and elegant all at the same time. The impacts are conditional, flexible, adaptive and open ended.
The inner structures of existence code for secondary and extended qualities more than they do for form. Of all the billions of humans, we are all individuals because the instructions of our existence call for tendencies, potentials, opportunities, not for absolutes or for perfect replicates. The cascade and orchestration of information and energy wells up from within us in one form and transforms through expanded communications of myriads of systems, unpredicted, yet "planned". Arms, legs, heart, mind, ...and the open option space to perform as no one else has ever performed "exactly". No atom "codes" for you. No single molecule "codes" for you. No single organelle or cell "codes" for you. What "codes" for you are the interactions and relations and encounters which those atoms, and molecules and cells have. Behaviors impacting behaviors producing behaviors being affected by behaviors.
This is the quality and the reality of Complexity that is our universe. Relationships are the superior aspect of existence and being. And they can be profound, so awe inspiringly, profound. Integrity of one, integrity of all.

References from "Braindance"
10. To give credit where it is due, Tobias first described the O/M sinus system in robust australopithecines in his classic monograph on Zinjanthropus. See P.V.Tobias, Olduvai Gorge, vol 2. (New York: Cambridge University Press, 1967). Holloway was the first to note that an O/M sinus system is also present in the Hadar fossil material in R.L.Holloway, "The endocast of the Omo juvenile L338y-6 hominid specimen, " American Journal of Physical Anthropology 54 (1981):109-18.
11. H. Browning, "the confluence of dural venous sinuses," American Journal of Anatomy 93 (1953):307-29.
![]()
THE INTEGRITY PAPERS (links to CEPTUAL READINGS)
GENRE WORKS (other writers)
POETICS
MINDWAYS (links to GLOBAL THINKERS)